Анатомия и эмбриология не могут доказать дарвинизм

АНАТОМИЯ И ЭМБРИОЛОГИЯ НЕ МОГУТ ДОКАЗАТЬ ДАРВИНИЗМ
Еще хуже дела обстоят в области «эволюционной» сравнительной анатомии, морфологии и эмбриологии. Потому что из этих фактов уже не то что не «вытащить» конкретную силу (или механизм), которая привела к эволюционным изменениям, но даже нельзя утверждать, что сам факт изменения таксонов (их происхождение друг из друга) имел место.
Действительно, ну и что, что (допустим) эмбрионы разных живых существ похожи друг на друга? Троллейбусы и автобусы - тоже похожи. Однако отсюда не следует, что троллейбусы произошли от автобусов в ходе эволюции, да еще и по механизму «случайные мутации+естественный отбор». Наоборот, мы точно знаем, что троллейбусы и автобусы возникли независимо. Если здесь и была какая-то зависимость, то только в головах у изобретателей и конструкторов этих троллейбусов и автобусов, которые использовали сходные планы при создании этих машин.
Тем более забавно звучит опора на морфологическое сходство (как доказательство чего-то там)… в рамках самой же теории эволюции, которая свободно оперирует понятием конвергенция форм. Конвергенция форм, в переводе с «эволюционного языка» на русский, как раз и означает именно сходство, но сходство таких видов, которые заведомо не являются друг другу родственниками. Например, теория эволюции предлагает нам поверить, что очень серьезное сходство между обыкновенным волком и австралийским сумчатым волком - это не более чем случайное эволюционное совпадение. А на самом деле, последний общий предок этих животных жил аж 160 млн. лет назад (см. ниже).
Еще теория эволюции уверяет нас (опираясь на молекулярно-генетические данные), что явное сходство ястребиных и соколиных птиц (Рис. 25) - это тоже случайное эволюционное совпадение. А на самом деле, эти группы птиц довольно далеки друг от друга. Соколиные генетически сгруппировались с воробьиными и попугаями, в то время как ястребиные оказались в заметном отрыве (Hackett et al., 2008). Отсюда следует, что к своему хищническому образу жизни соколиные и ястребиные пришли независимыми эволюционными путями (Hackett et al., 2008).

Рисунок 25. На этих фотографиях изображена совсем не одна и та же птица в разных ракурсах. На левой фотографии - полосатый лесной сокол (Micrastur ruficollis). На правой фотографии - ястреб-перепелятник (Accipiter nisus). Получается (согласно молекулярно-генетическим данным), что птица на левой фотографии ближе к попугаям и к воробьям, чем к птице на правой фотографии.
Ну и как после таких кульбитов с «конвергенцией форм» у теории эволюции хватает смелости уверять, что то же самое морфологическое сходство в других случаях доказывает эволюционное происхождение биологических таксонов друг из друга?
Получается очень забавная ситуация, которую можно описать таким образом:
- Морфологическое сходство организмов (морфологическое, анатомическое, эмбриологическое) доказывает эволюционное происхождение биологических таксонов друг из друга... кроме тех случаев, когда морфологическое сходство организмов не доказывает эволюционное происхождение таксонов друг из друга (сходство имеют такие биологические таксоны, которые друг из друга не происходили)...
Это классический пример таких «предсказаний» современной теории эволюции, которые автоматически делают из этой теории просто гения правильных предсказаний. Действительно, если мы скажем, что:
- Согласно нашей теории, в четверг должен пойти дождь или дождь пойти не должен...
То наша теория окажется наисильнейшей. Ведь что бы теперь ни случилось в четверг - мы всё равно правильно предсказали это событие!
Дело многократно осложняется еще и тем, что на самом деле, уже упомянутые нами эмбрионы - во многих отношениях не похожи друг на друга. Эмбрионы похожи только на рисунках Геккеля, который, как известно, чересчур увлёкся их рисованием, пририсовав им то, чего не было у оригиналов (или наоборот, недорисовав). Чтобы понять, насколько «стилизованные» рисунки Геккеля отличаются от оригиналов, можно посмотреть на рисунок 26:78
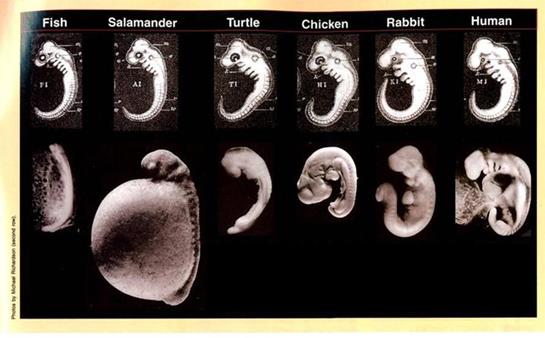
Рисунок 26. О том, как по-разному представляют себе развитие эмбрионов верующие дарвинисты и креационисты. Рисунок взят с креационного сайта «Разумный замысел».79
Верхний ряд: «Стилизованные» изображения разных эмбрионов (на ранней, «хвостовой» стадии), выполненные Эрнстом Геккелем. Рисунки эмбрионов «стилизованы» таким образом, чтобы бросалось в глаза их максимальное сходство - все эмбрионы приведены к сходному размеру; у них ярко прорисованы пресловутые «жаберные дуги»; «жаберные дуги» выглядят однотипно, и т.п. В результате создается ощущение, что эмбрионы разных классов позвоночных чуть ли не отштампованы друг от друга.
Нижний ряд: Фотографии эмбрионов некоторых видов животных (соответствующих классов), взятые для сравнения из разных рисунков работы Ричардсона (Richardson et al., 1997). Очевидно, что на самом деле, эмбрионы позвоночных на «хвостовой» стадии развития далеко не так похожи друг на друга, как это виделось Геккелю. Слева направо: рыба Salmo salar, саламандра Cryptobranchus allegheniensis, черепаха Emys orbicularis, курица Gallus gallus, кролик Oryctolagus cuniculus, человек Homo sapiens.
То есть, в реальности картина гораздо сложнее – у эмбрионов разных животных имеются как «похожести», так и «непохожести». И объяснение всех эмбриональных подробностей именно с помощью общего происхождения наталкивается на множество конкретных трудностей и несоответствий. Например, у эмбрионов разных позвоночных имеются серьезные несоответствия во времени формировании сердца и артерий, зачатков конечностей и даже «жаберных дуг».80 Но я не буду долго рассуждать на эту тему (пусть это делают профессиональные эмбриологи). Я приведу лишь один конкретный пример, который на мой взгляд, является весьма забавным.
Как нам известно еще со школьной скамьи,81 у эмбрионов всех позвоночных животных на ранних стадиях развития формируются так называемые «жаберные дуги». Это обстоятельство будто бы говорит нам о том, что все мы произошли от каких-то древних рыб. На рисунках Геккеля ярко выраженные «жаберные дуги» прямо-таки бросаются в глаза у всех эмбрионов.
На самом деле, это никакие не «жаберные дуги», а так называемые фарингеальные складки (или фарингеальные дуги, или висцеральные дуги), из которых у эмбрионов разных животных потом развиваются разные органы. Например, у человека из фарингеальных складок развиваются кости верхней и нижней челюсти, нёбная и скуловая кости, косточки слухового аппарата, хрящи гортани и трахеи и др. И никакими жабрами здесь и не пахнет.
«Жаберными дугами» фарингеальные складки эмбрионов называют только глубоко верующие дарвинисты. Ведь они искренне верят, что фарингеальные складки эмбрионов наземных позвоночных – это наследие «рыбьего» эволюционного прошлого. Нормальные же люди фарингеальные складки «жаберными дугами» не называют. Потому что на самом деле, эмбрионы наземных позвоночных никакими жабрами не дышат. Такое зрелище (дышащий жабрами эмбрион человека в утробе матери) может привидеться эмбриологу только в кошмарном сне. От которого эмбриолог проснётся в холодном поту.
На столь же серьезных основаниях можно смело называть зачатки конечностей у человеческого эмбриона – плавниками. Или, например, ластами. Или, при желании, «зачатками крыльев». Действительно, обозвать «зачатками крыльев» формирующиеся конечности человека, с научной точки зрения – полностью аналогично обзыванию фарингеальных складок «жаберными дугами».
А почему тогда не назвать нос человека – зачатком хобота? Ведь у слонов на эмбриональной стадии развития из этого места образуется именно хобот.
Здесь всё зависит от того, кого мы предпочтём записать в свои эволюционные предки или потомки. Если мы вдруг решим, что слоны произошли от людей, тогда формирующиеся на стадии эмбриона носы слонов и людей можно будет рассматривать в качестве эмбриологического доказательства происхождения слонов из людей. Точно так же, как фарингеальные складки эмбриона человека «доказывают» его происхождение от рыбы. Понятно, что всё это – бред, который способны на полном серьёзе обсуждать только самые верующие дарвинисты. Причем особенную пикантность этой ситуации придаёт тот факт, что у некоторых рыб (например, у данио) «жаберные дуги» на «хвостовой» стадии развития эмбриона… еще не образуются (Richardson et al., 1997). В то время как у наземных позвоночных они на этой стадии уже имеются. То есть, получается, что фарингеальные складки у эмбриона человека являются «доказательством» его происхождения от рыб… в то время как у самих рыб на этой же стадии еще нет никаких фарингеальных складок.82
И чтобы совсем уже закончить эту «жгущую» тему про «жаберные дуги», предлагаю посмотреть на соответствующий рисунок, где показаны фарингеальные складки эмбрионов разных видов животных на «хвостовой» стадии развития (Рис. 27):

Рисунок 27. Вид головы и шейного отдела у некоторых эмбрионов позвоночных животных: а – морская минога (Petromyzon marinus); b – глазчатый электрический скат (Torpedo ocellata); с – стерлядь (Acipenser ruthenus), d – листовая лягушка коки (Eleutherodactylus coqui); e - европейская болотная черепаха (Emys orbicularis); f – домашняя курица (Gallus gallus); g – лисовидный поссум (Trichosurus vulpecula); h – домашняя кошка (Felis catus). Взято из работы (Richardson et al., 1997).
Как видно из рисунка, фарингеальные складки (у тех эмбрионов, у которых они вообще имеются) различаются между собой весьма заметно. Эта реальная картина входит в диссонанс с теми идиллическими рисунками, которые нарисовал нам в своё время Эрнст Геккель. Там «жаберные дуги» разных эмбрионов повторяли друг друга чуть ли не под копирку (см. рис. выше). Впрочем, стоит ли удивляться? Ведь мощь теории эволюции очень велика. Если желание «доказать эволюцию» становится особенно сильным, то это желание, видимо, начинает влиять на объективную реальность. И под суровым взглядом верующего дарвиниста фарингеальные складки разных эмбрионов начинают выстраиваться в однотипные и аккуратные рядочки «жаберных щелей».
Вообще, «эмбриологическими доказательствами эволюции» дарвиниста можно тихо троллить.83
Например, как оказалось, у некоторых рыбьих эмбрионов84 на определенном этапе развития имеется стадия, когда в формирующихся конечностях (будущих плавниках) начинают работать именно те самые гены, которые ответственны за образование у четвероногих – кисти и пальцев (именно кисти и пальцев, а не конечности в целом). Причем эти гены начинают работать именно в том месте и в то самое время, где у наземных позвоночных закладываются и формируются кисть и пальцы. В рамках теории эволюции это открытие стало большим сюрпризом. Никто не ожидал, что подобная стадия может обнаружиться и у рыб тоже. Ведь согласно современной теории эволюции, рыбы никогда никаких кистей и пальцев не имели. Эти части конечности считались более поздним изобретением эволюции (связанным с выходом животных на сушу).
Тем более что в предыдущих исследованиях у рыбки данио этой особой (второй) фазы работы Hox-генов (которая у наземных позвоночных ответственна за формирование кисти и пальцев), обнаружено не было.85Поэтому было естественно предположить, что вторая фаза работы этих генов явилась новоприобретением нашей любимой эволюции, когда рыбы стали «выходить на сушу».
Однако реальность оказалась сложнее простых эволюционных схем.
Когда работу этих же генов решили посмотреть у рыб-веслоносов (Davis et al., 2007) и еще у акул (Freitas et al., 2007), то у эмбрионов этих рыб вдруг обнаружилась именно та самая (вторая) фаза экспрессии тех генов, которые у наземных позвоночных отвечают за формирование кистей и пальцев. Обнаружилась именно в том месте и в то время, где и положено.86
Однако мы знаем, что у акул и осетров никаких пальцев не образуется.87Видимо, вторая фаза экспрессии обсуждаемых генов у этих рыб быстро затухает или (может быть) блокируется? А у наземных позвоночных продолжается. В результате чего у них и образуются пальцы.
Объяснить этот интересный факт можно разными способами. Проще всего это объяснить следствием общего дизайна всех позвоночных. Можно предположить, что все позвоночные животные производятся из одной сходной «универсальной заготовки» (со сходными генами), которую эмбрионы позвоночных образуют на определенной стадии развития. А дальше за счет того, что имеющиеся общие гены начинают включаться и выключаться в разное время, работать с разной продолжительностью и интенсивностью - в итоге получаются весьма разные животные.
Такую идею (об «универсальной заготовке» всех позвоночных) подтверждает еще один интересный факт. Дело в том, что эмбрионы разных классов позвоночных, на самом деле, приходят к этой «общей стадии» весьма разными путями (начиная от разных типов дробления яйца). Такая модель развития эмбрионов позвоночных получила название модели «песочных часов» (Рис. 28):

Рисунок 28. Так называемая «модель песочных часов». Модель «песочных часов» призвана отразить тот факт, что эмбрионы позвоночных животных сильнее всего различаются между собой на ранних стадиях своего развития (начиная от дробления яйца по разному типу), и на поздних стадиях развития. А вот где-то примерно «посерединке» эмбрионы позвоночных образуют сходную стадию. Рисунок выполнен на основе соответствующего рисунка из работы (Richardson et al., 1997), с небольшими уточнениями.
Понятно, что сходная стадия, почему-то образующаяся где-то «посерединке» эмбрионального развития из весьма непохожих исходных состояний, прямо-таки напрашивается на то, чтобы считать эту стадию именно «универсальной заготовкой» (из которой потом, может быть, просто удобно программировать дальнейшее развитие животных разных классов).
Интересно, что примерно такой же принцип развития эмбрионов (по модели «песочных часов») был обнаружен и у некоторых насекомых, представителей отряда двукрылых (Kalinka et al., 2010). Причем в этом случае сверялись не морфологические признаки, а работа разных генов на разных стадиях развития эмбриона. Тем не менее, получились тоже «песочные часы». Более того, примерно то же самое было обнаружено еще и у растений (Quint et al., 2012).
Итак, проще всего такие факты объясняются в рамках представлений о едином дизайне эмбрионального развития (тех или иных биологических таксонов).
Но можно объяснить озвученные факты и в рамках эволюционных представлений об общем предке. Например, развитие эмбрионов разных классов животных по модели «песочных часов» можно объяснить, сделав предположение, что интригующая «сходная стадия» всех позвоночных является наиболее ответственной и уязвимой (в смысле фатальных ошибок развития). Поэтому в ходе эволюции эта стадия оказалась наиболее консервативной и лучше всего сохранялась. А более ранние и более поздние стадии развития эмбрионов такого консерватизма уже не требовали. И поэтому изменялись более свободно.
Ну а в отношении выше озвученных «генов кисти и пальцев» у рыб - тут можно предположить, что вторая фаза работы соответствующих Hox-генов у хрящевых и хрящекостных рыб – исходно была нужна им для формирования каких-то других вещей в плавниках (а не кистей и пальцев). Однако в ходе дальнейшей эволюции эта фаза экспрессии перестроилась таким образом, что начала формировать кисть и пальцы появившимся наземным позвоночным. Примерно к такому выводу (об исходной древности программы развития дистальных частей конечностей) склонились сами авторы тех работ, в которых были открыты эти факты.
Итак, мы видим, что эмбриональное сходство (или различие) можно свободно трактовать как в рамках эволюционной концепции, так и в рамках разумного дизайна. Поэтому никакими «доказательствами эволюции» подобные факты являться не могут.
Более того, можно начать «объяснять» эти же факты в эволюционном свете, но при этом развернув их в противоположную сторону. Например, можно начать упирать на то, что вторая фаза работы соответствующих Hox-генов у хрящевых и хрящекостных рыб (наблюдаемая сегодня), скорее всего, является исходным состоянием когда-то исправно работавшей системы развития кистей и пальцев у этих животных. И в полном соответствии с «эмбриологическими доказательствами эволюции», заявить, что хрящевые и хрящекостные рыбы произошли от наземных четвероногих животных (а не наоборот). О чем и свидетельствуют явные «остатки» генетической программы формирования кисти и пальцев у эмбрионов озвученных рыб. Таким образом, Вы можете спокойно заявить, что акулы и осетры, скорее всего, произошли от каких-то наземных позвоночных. Например, от пингвинов.
Тем более что и знаменитые «жаберные дуги» у (некоторых) рыб развиваются на более поздних эмбриональных стадиях, чем у наземных позвоночных (см. выше). Отсюда, в полном соответствии с логикой «эмбриологических доказательств эволюции» следует, что жабры рыб являются более поздним эволюционным приобретением.
Это будет хорошим троллингом дарвиниста, верующего в «эмбриологические доказательства эволюции». Вы можете спросить такого дарвиниста – как он сам считает – является ли обнаруженная экспрессия «генов кисти и пальцев» у рыб доказательством того, что акулы произошли от наземных четвероногих животных?
С вероятностью 99% верующий дарвинист ответит Вам так:
- Нет, не является! Эти гены не свидетельствуют о том, что у рыб когда-то были пальцы! Просто эти гены нужны рыбам для чего-то другого!
После получения подобного ответа задайте Вашему дарвинисту следующий вопрос:
- Ну и для чего же нужны эти гены («пальцев») рыбам?
На этот вопрос дарвинист ответит примерно следующее:
- Я не знаю, но для чего-то, конечно же, нужны.
В общем, после того, как Вы дождётесь, что дарвинист произнесёт примерно следующее:
- Они (эти гены) нужны для чего-то другого, а не являются доказательством эволюции рыб из наземных позвоночных…
Вот после этого Вы сможете нанести верующему дарвинисту нокаутирующий удар. Примерно таким образом:
- А чего это Вы так свободно перепрыгиваете с одного объяснения на другое? Когда Вам надо, Вы списываете черты сходства эмбрионов на «общее происхождение». Например, Вы почему-то называете фарингеальные складки человеческого эмбриона… «жаберными дугами» (!), хотя это никакие не «жаберные дуги», а фарингеальные складки. Ведь никакие жабры из них не развиваются, а развиваются совершенно другие органы. То есть, это именно та ситуация, когда фарингеальные складки «нужны для чего-то другого», а не для формирования жабр. Причём даже известно, для чего.88
Но в другом случае Вы почему-то не называете работающие у эмбрионов рыб гены «кистей и пальцев» - генами «кистей и пальцев». И не считаете их доказательством того, что рыбы произошли от наземных четвероногих животных. А считаете, что эти гены «просто нужны для чего-то другого». Хотя даже понятия не имеете, для чего именно.
Разве Вы не видите, что таким образом демонстрируете совершенно откровенные двойные стандарты в Ваших «доказательствах эволюции» с помощью «похожести эмбрионов»? И какова после этого цена подобным «доказательствам эволюции»?
В 99% случаев именно на этой эффектной ноте можно спокойно закончить дискуссию на эту тему.
Однако теоретически существует возможность, что Вам (вдруг) попадётся какой-нибудь особенно дотошный дарвинист, который твердо решит докопаться до истины в этом вопросе - узнать, для чего же всё-таки нужны рыбам «гены кистей и пальцев». Такой дарвинист может начать поиск оригинальных научных исследований, где этот вопрос изучался. Лично мне настолько дотошные дарвинисты еще ни разу не попадались. Обычно они оперируют фактами из того списка литературы, который я уже озвучил в самом начале этой книги (учебники биологии, статьи из википедии, книжки Докинза или Маркова, в крайнем случае, какие-нибудь научно-популярные статьи на эволюционную тематику).
Тем не менее, теоретически не исключена возможность, что Вам попадется настолько дотошный дарвинист, что его не испугает даже чтение оригинальных научных публикаций. В этом случае он сможет заявить Вам (опираясь, например, на работу Freitas et al., 2014), что так называемый апикальный эктодермальный гребень, действительно, образуется в дистальной части формирующихся конечностей как у эмбрионов рыб, так и у наземных позвоночных. И действительно, у наземных позвоночных этот апикальный гребень просто продолжает развиваться дальше - вплоть до формирования дистальной части конечности наземного позвоночного (кистей, стоп и пальцев). Однако и у рыб апикальный эктодермальный гребень тоже не«рассасывается» (следовательно, не является рудиментом). Он превращается позднее в так называемую апикальную эктодермальную складку (Freitas et al., 2014),89 которая в свою очередь (наверное) превращается дальше в какие-то дистальные части рыбьего плавника.
Если дарвинист всё-таки дочитается до всего этого, то Вам надо будет немного продолжить Вашу аргументацию. Вы можете ответить дарвинисту примерно следующее:
- В том то и дело. Апикальный эктодермальный гребень имеется и у рыб, и у наземных позвоночных. Но у наземных позвоночных этот гребень потом просто продолжает своё развитие, вплоть до формирования пальцев. А у рыб этот же (исходный) гребень сначала превращается в апикальную эктодермальную складку, из которой лишь потом формируются (наверное) какие-то дистальные части плавника. То есть, у рыб здесь имеется дополнительная стадия превращения. И отсутствие этой дополнительной стадии – является условием для формирования конечности именно по типу наземных позвоночных (с кистями и пальцами).
Из этого факта как раз и получается (если следовать логике Геккеля), что исходным эволюционным состоянием был именно апикальный эктодермальный гребень, и следовательно, кисть и пальцы наземных позвоночных. А вот апикальная эктодермальная складка – это уже более позднее эволюционное производное от этого гребня. Следовательно, дистальные части рыбьего плавника – это тоже более позднее эволюционное образование, которое возникло уже на основе зачатка кистей и пальцев наземных позвоночных (который преобразовался соответствующим образом в ходе эволюции). Таким образом, если следовать логике Геккеля – рыбы действительно произошли от наземных позвоночных. Во всяком случае, озвученные факты указывают именно на такой эволюционный сценарий.
Примечания.
78. Многие верующие дарвинисты нередко защищают Эрнста Геккеля, доказывая, что Геккель не фальсифицировал рисунки (с целью «доказать эволюцию»), а просто «схематизировал их для лучшего понимания». Понятно, что в таком «ракурсе» становится трудно отделить сознательную фальсификацию от «сознательной схематизации». Однако некоторые манипуляции Геккеля, которые он проделал с рисунками эмбрионов, всё-таки заставляют считать эти манипуляции больше фальсификацией, чем «схематизацией». Вот что пишут авторы соответствующей работы по этому поводу (Richardson & Keuck, 2002):
«The schematisation defence is not easy to reconcile with the observation that limb buds were removed by Haeckel from some early embryos (Richardson and Keuck, 2001). Indeed, one could argue that such selective schematisation is no different from fraud»
Перевод: «Схематизационной защите» нелегко смириться с наблюдением, что зачатки конечностей у некоторых ранних эмбрионов были удалены Геккелем. На самом деле можно утверждать, что такая селективная «схематизация» не отличается от мошенничества.
То есть, зачатки конечностей, которые у некоторых эмбрионов, видимо, появляются слишком рано (по мнению Геккеля) - были удалены Геккелем. Наверное для того, чтобы этот факт не «вводил в искушение» возможных читателей. Ведь чересчур раннее появление зачатков конечностей может заставить засомневаться в том, что наземные позвоночные произошли от рыб.
79. http://www.origins.org.ua/page.php?id_story=1315 взято 5.09.2014
80. Сердце у высших позвоночных (амниот) формируется раньше, чем у рыб. У крыс система аорты начинает просматриваться уже на стадии фарингулы. У сумчатых передние конечности развиваются гораздо раньше, чем у других наземных позвоночных и т.п. (Richardson et al., 1997).
81. Из учебника общей биологии для средней школы с рисунками Геккеля прямо на обложке.
82. Во всяком случае, у данио рерио (Danio rerio).
83. Троллинг в современной культуре интернет-общения – это издевательство над собеседником (или собеседниками). Может выражаться в завуалированной, тонкой форме, или, наоборот, в откровенно грубой и т.п. Происходит от слова «тролль».
84. Из таксонов хрящевых и хрящекостных рыб.
85. Отряд карпообразные, инфракласс костистые рыбы.
86. Еще позднее оказалось, что и у рыбки данио соответствующая фаза экспрессии этих генов тоже, вроде бы, всё-таки имеется (Ahn & Ho, 2008).
87. Веслоносы, у которых была обнаружена обсуждаемая вторая стадия экспрессии соответствующих Hox-генов (Davis et al., 2007), принадлежат к отряду осетрообразных (подкласс хрящекостные рыбы).
88. Из фарингеальных складок млекопитающих образуются кости верхней и нижней челюсти, нёбная и скуловая кости, косточки слухового аппарата, хрящи гортани и трахеи и др.
89. «In fish the AER elongates after fin initiation forming an apical ectodermal fold (AEF) where the dermoskeleton differentiates (Fig.3A,B; Thorogood, 1991). In tetrapods, however, the AER is never reverted into an AEF and persists until the formation of digits (Fig.3С; Mariani and Martin, 2003)» (Freitas et al., 2014).
Илья Рухленко, Что ответить дарвинисту


